 De fleste har interesse for slekta si bakover. Noen kan følge den i flere hundre år. Men de fleste må si seg tilfreds med å lese historiebøker og forestille seg hvordan forfedrene levde for hundrevis av år siden (jfr. TV-serien Anno) Hvor langt tilbake kan vi få informasjon om slekta vår? Bibellesere vet at menneskeslekten kan føres til Noah og kona Naamah, og at forut for det var det noen første eksemplarer av menneskeslekten (Adam og Eva).
De fleste har interesse for slekta si bakover. Noen kan følge den i flere hundre år. Men de fleste må si seg tilfreds med å lese historiebøker og forestille seg hvordan forfedrene levde for hundrevis av år siden (jfr. TV-serien Anno) Hvor langt tilbake kan vi få informasjon om slekta vår? Bibellesere vet at menneskeslekten kan føres til Noah og kona Naamah, og at forut for det var det noen første eksemplarer av menneskeslekten (Adam og Eva). Mange vitenskapsfolk, biologer og andre, mener at vi må gå mye lengre bakover enn det. Langt bakover i en fjern fortid ser de våre stamfedre, som ape-lignende forfedre. Nå har vi tidligere sett at tidsrommet mellom slektene er for kort til at tilfeldig neo-darwinistisk utvikling skal kunne gjøre rede for overgangen. Lengre bakover skulle vår stamfar være et slags insektetende pattedyr. Enda lengre bakover på vårt familietre, skal det ha vært fisk som var stamfaren (noe vi også har kommentert tidligere).
 Et hoveddogme i NDT er at mutasjoner ikke i det hele er relatert til organismenes behov. Teorien er basert på tilfeldige genetiske endringer. Om evolusjon ble funnet å relatere til miljømessige endringer eller organismens behov, så ville det motsi NDT. Evolusjonister har stresset igjen og igjen at variasjoner som evolusjonen stammer fra er tilfeldige i så henseende.
Et hoveddogme i NDT er at mutasjoner ikke i det hele er relatert til organismenes behov. Teorien er basert på tilfeldige genetiske endringer. Om evolusjon ble funnet å relatere til miljømessige endringer eller organismens behov, så ville det motsi NDT. Evolusjonister har stresset igjen og igjen at variasjoner som evolusjonen stammer fra er tilfeldige i så henseende. 3. Nøytrale variasjoner har ingen effekt på fruktbarheten og utgjør ingen forskjell i forhold til utviklingen. De påvirker ikke organismens evne til å overleve for å formere seg. Øyefarge i mennesker synes f.eks. å være en slik nøytral variasjon.
3. Nøytrale variasjoner har ingen effekt på fruktbarheten og utgjør ingen forskjell i forhold til utviklingen. De påvirker ikke organismens evne til å overleve for å formere seg. Øyefarge i mennesker synes f.eks. å være en slik nøytral variasjon. Anta f.eks. at et dyr i en populasjon med stabil størrelse, i gjennomsnitt produserer 5 avkom i sin reproduktive livsfase. Siden populasjonen er av stabil størrelse, vil bare ett av 5 av disse overleve for å reprodusere seg. Det innebærer at sjansene bare er 20% for at et spesielt dyr ved fødselen vil overleve til å reprodusere seg. Om det overlever eller ikke, beror i hovedsak på ytre omstendigheter. Egne evner kan ha lite å si. En mutant med en SV på 0,1% vil ha en sjanse på 20,02% i stedet for 20% på å overleve. Sannsynligheten for at den vil overleve 1.generasjon er 0,2002. Sannsynligheten for at den vil overleve 2. generasjon er 0,1347 (Fishers beregning). Muligheten for å overleve 3.generasjon er enda mindre. I standardpopulasjonen er sannsynligheten for å overleve 1.generasjon 0,2, de førse to generasjoner: 0,1345 etc. For å få fram en signifikant forskjell mellom enkelt-mutanter og andre, må 4 desimaler benyttes.
Anta f.eks. at et dyr i en populasjon med stabil størrelse, i gjennomsnitt produserer 5 avkom i sin reproduktive livsfase. Siden populasjonen er av stabil størrelse, vil bare ett av 5 av disse overleve for å reprodusere seg. Det innebærer at sjansene bare er 20% for at et spesielt dyr ved fødselen vil overleve til å reprodusere seg. Om det overlever eller ikke, beror i hovedsak på ytre omstendigheter. Egne evner kan ha lite å si. En mutant med en SV på 0,1% vil ha en sjanse på 20,02% i stedet for 20% på å overleve. Sannsynligheten for at den vil overleve 1.generasjon er 0,2002. Sannsynligheten for at den vil overleve 2. generasjon er 0,1347 (Fishers beregning). Muligheten for å overleve 3.generasjon er enda mindre. I standardpopulasjonen er sannsynligheten for å overleve 1.generasjon 0,2, de førse to generasjoner: 0,1345 etc. For å få fram en signifikant forskjell mellom enkelt-mutanter og andre, må 4 desimaler benyttes.
 En kan sammenligne punktmutasjoner, endringer i en enkelt DNA-nukleotide, med tilfeldige endringer i bokstaver i en tekst. Anta at en (post-moderne) forfatter ville prøve å få til forbedringer i en novelle ved tilfeldig å endre bokstaver i den. Om han fulgte evolusjonens regler, så måtte han velge enten den endrede teksten eller original-teksten, hele pakken eller ingenting. Han kunne ikke velge mellom enkeltendringer. Om en skulle holde seg etter neo-darwinismen, måtte mutasjonsraten være lav, ca. én bokstav i novellen pr. forsøk. Om den ble for høy, ville det bli mange endringer og de fleste ville være negative. Hver gang han fikk en forbedring, var det sannsynlig at det kom en skade -så han måtte forkaste den endrede versjonen. Anta han fikk en bokstavs endring i novellen. Han kunne f.eks. få ordet fest til å endre seg til ett hvilken som helst av ordene: gest, hest, rest.
En kan sammenligne punktmutasjoner, endringer i en enkelt DNA-nukleotide, med tilfeldige endringer i bokstaver i en tekst. Anta at en (post-moderne) forfatter ville prøve å få til forbedringer i en novelle ved tilfeldig å endre bokstaver i den. Om han fulgte evolusjonens regler, så måtte han velge enten den endrede teksten eller original-teksten, hele pakken eller ingenting. Han kunne ikke velge mellom enkeltendringer. Om en skulle holde seg etter neo-darwinismen, måtte mutasjonsraten være lav, ca. én bokstav i novellen pr. forsøk. Om den ble for høy, ville det bli mange endringer og de fleste ville være negative. Hver gang han fikk en forbedring, var det sannsynlig at det kom en skade -så han måtte forkaste den endrede versjonen. Anta han fikk en bokstavs endring i novellen. Han kunne f.eks. få ordet fest til å endre seg til ett hvilken som helst av ordene: gest, hest, rest. Fordi punktmutasjoner var de minst tenkelige endringer, så ble neo-darwinister glade for å kunne adoptere det som kilde til variasjon for sin teori. Men en kopieringsfeil er ikke ignorerbart liten, spesielt ikke når den må følges opp av korrelerte tilpasninger andre steder. De har ikke løsningen på den lille endringen, som Darwin benyttet til å vise hvordan øyet utviklet seg, faktisk vil virke. NDT sier endringer vil opptre tilfeldig, og at naturlig seleksjon på en måte ser på hver endring og bestemmer hvorvidt den vil droppe eller bevare den. På det viset vil en populasjon tilpasse seg til å endre seg, for å tilpasse seg miljøendringer. Men vi skal senere se at det ikke er noe bevis for at kompleksitet er bygd opp av prosesser beskrevet av NDT.
Fordi punktmutasjoner var de minst tenkelige endringer, så ble neo-darwinister glade for å kunne adoptere det som kilde til variasjon for sin teori. Men en kopieringsfeil er ikke ignorerbart liten, spesielt ikke når den må følges opp av korrelerte tilpasninger andre steder. De har ikke løsningen på den lille endringen, som Darwin benyttet til å vise hvordan øyet utviklet seg, faktisk vil virke. NDT sier endringer vil opptre tilfeldig, og at naturlig seleksjon på en måte ser på hver endring og bestemmer hvorvidt den vil droppe eller bevare den. På det viset vil en populasjon tilpasse seg til å endre seg, for å tilpasse seg miljøendringer. Men vi skal senere se at det ikke er noe bevis for at kompleksitet er bygd opp av prosesser beskrevet av NDT.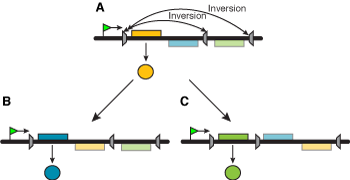 Noen evolusjonister har foreslått en annen måte å lagre mutasjoner for senere bruk. Et gen kan ha et stykke av det 'snudd rundt', fra en inversjon, eller ett ekstra stykke er satt inn (innsetting) som kan hindre genet fra å fungere. Om f.eks. et gen har et ekstra sett av basepar, så ville kodingssekvensen -måten kodoner (
Noen evolusjonister har foreslått en annen måte å lagre mutasjoner for senere bruk. Et gen kan ha et stykke av det 'snudd rundt', fra en inversjon, eller ett ekstra stykke er satt inn (innsetting) som kan hindre genet fra å fungere. Om f.eks. et gen har et ekstra sett av basepar, så ville kodingssekvensen -måten kodoner (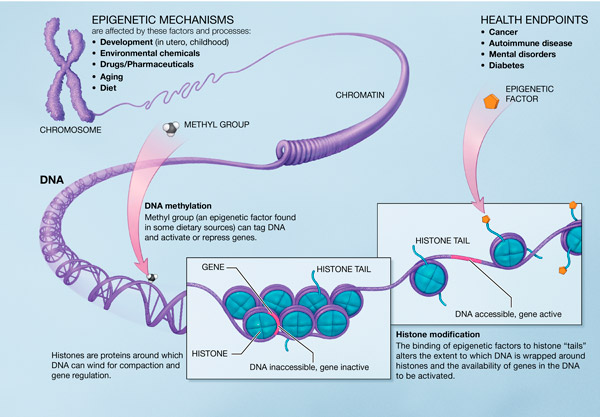 Det finnes noen alleler i en populasjon som har vist seg være nyttige i fortiden. Men en kan ikke forvente å finne nye gener liggende rundt omkring, som kan spille en nyttig rolle de aldri tidligere har spilt. Alleler som har vært nyttige i fortiden blir lagret i en populasjon og finnes i store antall, nærmere 1000 enn 1. Det kan være overlevninger fra noe som var en stor fraksjon av populasjonen, og som seleksjon ikke ennå har kvittet seg med. De kan bare være en inversjon eller innsetting unna å bli vekket opp til å fungere. Dess flere slike gener det er, desto lengre kan de ligge uvirksomme i populasjonen. Noen kan i og for seg være fungerende, men ikke være tilpasset for virksomhet nå. F.eks. kan epigenetiske brytere være slått av, inntil endringer i miljøet fører til at de slås på (
Det finnes noen alleler i en populasjon som har vist seg være nyttige i fortiden. Men en kan ikke forvente å finne nye gener liggende rundt omkring, som kan spille en nyttig rolle de aldri tidligere har spilt. Alleler som har vært nyttige i fortiden blir lagret i en populasjon og finnes i store antall, nærmere 1000 enn 1. Det kan være overlevninger fra noe som var en stor fraksjon av populasjonen, og som seleksjon ikke ennå har kvittet seg med. De kan bare være en inversjon eller innsetting unna å bli vekket opp til å fungere. Dess flere slike gener det er, desto lengre kan de ligge uvirksomme i populasjonen. Noen kan i og for seg være fungerende, men ikke være tilpasset for virksomhet nå. F.eks. kan epigenetiske brytere være slått av, inntil endringer i miljøet fører til at de slås på ( Om det er slik at endring i ett bits informasjon fører til mye ny kompleksitet, så måtte en mistenke at kompleksiteten var der allerede i utgangspunktet. Den rette mutasjon i ett regulerende gen, kan slå på en hel rad av latente gener. I prinsippet kan en enkelt mutasjon i regulerende gen påvirke et helt kompleks av aktivitet. En mutasjon som reverserer effekten av en tidligere, kunne i ett nu gjenopplive komplekse funksjoner som tidligere var slått av. Men siden en enkelt mutasjon ikke kan endre mer enn en bit informasjon, skjønner vi at kompleksiteten må ha vært tilstede i genomet tidligere. Mutasjonen måtte ha slått PÅ noe som har vært et eksisterende, men inaktivt system. En mutasjon som slår PÅ en hvilende funksjon, er ikke det som kreves for NDT-versjon av utvikling. Hvorfra skulle de hvilende genene komme? Hvordan utviklet de seg? En kan få evolusjon på dette viset, men det er ikke neo-darwinistisk evolusjon. Vi kommer tilbake til dette. I visse tilfeller kan skje at mutasjoner mister informasjon og likevel tjener organismen i spesielle omstendigheter. De fleste av disse mutasjonene mister informasjon fordi de deaktiverer repressor gener.
Om det er slik at endring i ett bits informasjon fører til mye ny kompleksitet, så måtte en mistenke at kompleksiteten var der allerede i utgangspunktet. Den rette mutasjon i ett regulerende gen, kan slå på en hel rad av latente gener. I prinsippet kan en enkelt mutasjon i regulerende gen påvirke et helt kompleks av aktivitet. En mutasjon som reverserer effekten av en tidligere, kunne i ett nu gjenopplive komplekse funksjoner som tidligere var slått av. Men siden en enkelt mutasjon ikke kan endre mer enn en bit informasjon, skjønner vi at kompleksiteten må ha vært tilstede i genomet tidligere. Mutasjonen måtte ha slått PÅ noe som har vært et eksisterende, men inaktivt system. En mutasjon som slår PÅ en hvilende funksjon, er ikke det som kreves for NDT-versjon av utvikling. Hvorfra skulle de hvilende genene komme? Hvordan utviklet de seg? En kan få evolusjon på dette viset, men det er ikke neo-darwinistisk evolusjon. Vi kommer tilbake til dette. I visse tilfeller kan skje at mutasjoner mister informasjon og likevel tjener organismen i spesielle omstendigheter. De fleste av disse mutasjonene mister informasjon fordi de deaktiverer repressor gener. Sunn fornuft sier at den forbløffende kompleksiteten i livet ikke kan oppstå gjennom tilfeldige prosesser. Til tross for iherdige forsøk fra evolusjonister på å få naturlig seleksjon til å fremstå som ikke-tilfeldig, glemmer de hovedpoenget: Kan tilfeldige endringer gi naturlig seleksjon tilstrekkelig av de rette genene, så evolusjonen har noe å jobbe med? Darwin fungerte som en god skranke-advokat i å få en svak sak til å virke solid. W.R. Thompson, en høyt ansett biolog og medlem av Royal Society har sagt: " Darwin viste ikke i 'The Origin' at arter hadde oppstått ved naturlig seleksjon. Han viste bare på basis av visse fakta og formodninger, hvordan dette kunne ha skjedd, og som han hadde overbevist seg selv, var han i stand til å overbevise andre. (Thompson, 1963, s. xii).
Sunn fornuft sier at den forbløffende kompleksiteten i livet ikke kan oppstå gjennom tilfeldige prosesser. Til tross for iherdige forsøk fra evolusjonister på å få naturlig seleksjon til å fremstå som ikke-tilfeldig, glemmer de hovedpoenget: Kan tilfeldige endringer gi naturlig seleksjon tilstrekkelig av de rette genene, så evolusjonen har noe å jobbe med? Darwin fungerte som en god skranke-advokat i å få en svak sak til å virke solid. W.R. Thompson, en høyt ansett biolog og medlem av Royal Society har sagt: " Darwin viste ikke i 'The Origin' at arter hadde oppstått ved naturlig seleksjon. Han viste bare på basis av visse fakta og formodninger, hvordan dette kunne ha skjedd, og som han hadde overbevist seg selv, var han i stand til å overbevise andre. (Thompson, 1963, s. xii).